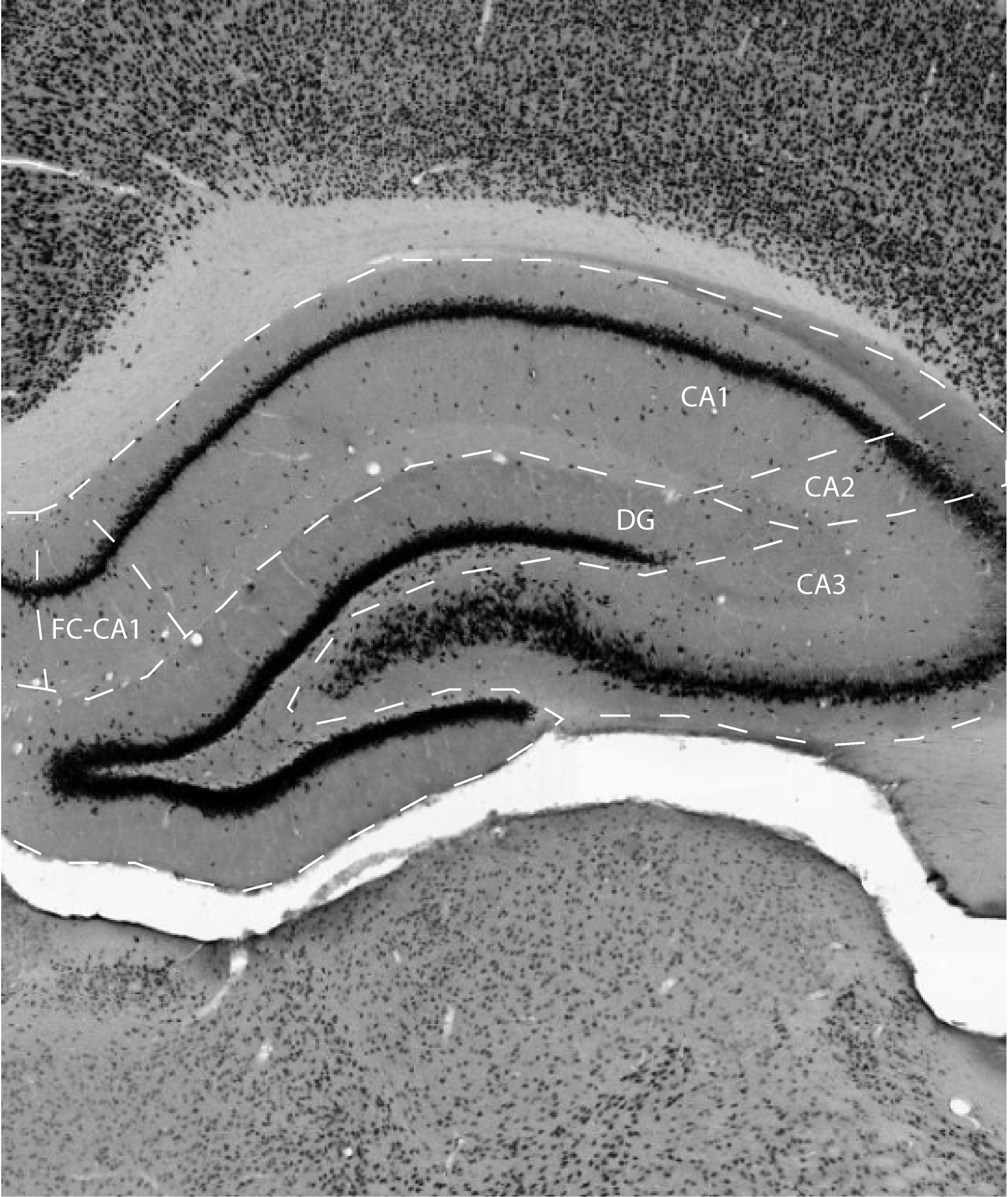
The Cornu Ammonis or Ammon's Horn (CA) also referred to as hippocampus proper, is characterized by a thin layer of densely packed pyramidal cells, enclosed by an outer plexiform layer and an inner polymorph layer, also called stratum oriens. The outer plexiform layer, which contains the apical dendrites of the pyramidal cells, is generally subdivided into a number of sublayers. These will be detailed in the descriptions of its three main subdivisions, area CA1, CA2, and CA3 (according to some accounts a field CA4 can be differentiated as well, but because this is ill-defined with respect to the hilus of the dentate gyrus, this practice will not be followed here (Witter, 2004). The CA field is bordered on one side by the dentate gyrus (DG); this side of CA is generally referred to as the proximal part. The opposite distal side is where CA meets the subiculum (SUB). The CA, and thus its constituting subfields, is a three-dimensionally complex structure such that its orientation is often difficult to grasp when seen in different planes of sectioning.
Area CA1 is part of the hippocampus proper, whereas the subiculum is
generally considered to represent a separate entity. However, both
areas share the layered composition which is typical for hippocampal
fields. This layered apprearance is most easily seen in Nissl, NeuN or
Timm-stained sections. In CA1, and less prominently in the subiculum, a
wide molecular layer is located between the pyramidal layer and the
hippocampal fissure. In the subiculum, this molecular layer can be
subdivided into a deeper portion that is continuous with the stratum
radiatum of CA1, and a superficial portion that is continuous with
stratum lacunosum-moleculare. Note that this subdivision is difficult
to detect by the markers used in this atlas. The superficial molecular
layer of the subiculum and its continuation in stratum lacunosum
moleculare of CA1 contain the perforant pathway fibers from the EC.
Moreover, afferents from other structures such as the nucleus reuniens
of the midline thalamus take a similar course and distribution.
CA1 can be easily recognised in Nissl and NeuN-stained sections due to
its layer of neatly aligned pyramidal cells. This feature distinguishes
CA1 from the subiculum, which dorsally is located between area CA1 and
the retrospenial cortex, and ventrally between area CA1 and pre- and
parasubiculum. The CA1/subiculum border is clearly marked by an abrupt
widening of the pyramidal cell layer. Moreover, in material stained for
parvalbumin or AChE, the pyramidal layer of CA1 is darkly stained,
whereas the pyramidal cell layer of the subiculum is more diffusely
stained, thus indicating a marked border between the two fields. In the
TIMM staining the border is also visible, but then with an unstained
CA1pyramidal layer, and a darkly stained cell layer in the proximal
part of the subiculum. Although the border may appear easy to
establish, it is in practice not possible to determine whether a
particular dendrite at the border belongs to a neurone in CA1 or in the
subiculum. This is caused by the oblique orientation of the CA1 /
subiculum border relative to the transverse axis. Therefore, cells (in
the stratum oriens, stratum radiatum, or lacunosum moleculare) close to
this border do not necessary extent their dendrites perpendicularly to
the orientation of stratum pyramidale.
References -> access a list of references
NIF Navigator (external link) -> search the Neuroscience Information Framework |
|